The use of physicochemical methods in the analysis of bioflavonoids and phenolcarboxylic acids in medicinal plant materials. Method for isolation and purification of phenolcarboxylic acids Nucleophilic substitution reaction with halogens
In addition, they are characterized by properties due to the presence in the molecule of both types of functional groups and a benzene ring.
Phenol acids are crystalline solids. Phenolic acids, which contain one phenolic hydroxyl, are relatively poorly soluble in cold water, but dissolves well in hot water and many organic solvents. With an increase in the number of phenolic hydroxyls, the solubility of phenolic acids increases.
Distribution in nature
Synthesis
Synthetic methods are often used to obtain phenolic acids. In particular, 2-hydroxybenzoic (salicylic) acid is extracted from sodium phenolate and carbon dioxide in autoclaves at 180 °C, followed by processing of the reaction product hydrochloric acid Kolbe synthesis:
C6H5ONa + С02 180 С → С6Н4 (ONa) COOH; C6H4 (ONa) COOH + HC1 → C6H4 (OH) COOH + NaCl
Chemical properties
Phenol acids simultaneously have the properties of carboxylic acids and phenols. In addition, they are characterized by properties due to the presence of both types of functional groups and a benzene ring in the molecule.
Heat decomposition
When heated, phenolic acids decompose to form phenolic compounds and carbon dioxide. For example, when heated, salicylic acid decomposes into phenol and carbon dioxide:
HOC6H4COOH → C6H5OH + CO2 salicylic acid phenol carbon dioxide
Esterification reaction (at the carboxyl group)
Phenol acids, like carboxylic acids, are capable of forming esters due to the presence of carboxyl and hydroxyl groups.
For example, the formation of salicylic acid ester - acetylsalicylic acid:
HOC6H4COOH + H3C-C(=O)-O-(O=)C-CH3 → C6H4(COOH)-O-CO-CH3 + CH3COOH salicylic acid acetic ester acetylsalicylic acid acetic acid
and the formation of gallic acid ester, one gallic acid molecule reacts with its carboxyl, the other with its phenyl hydroxyl. Chinese tannin is a glucoside of digallic acid and glucose.
Formation of salts
Phenol acids, like carboxylic acids, form salts. For example, the formation of sodium salicylate by the interaction of salicylic acid and sodium hydroxide:
HOC6H4COOH + NaOH → HOC6H4COONa + H2O salicylic acid sodium salicylate
Reaction with iron(III) chloride (at the phenolic group)
There is often a need to detect the presence of salicylic acid and other phenolic acids in canned foods. Then 2-3 ml of the test solution is placed in a test tube and a few drops of a 1% solution of iron (III) chloride are added. A purple color appears. Unlike phenol, it can also appear in alcohol solution acids. The color occurs as a result of the formation of complex salts during the interaction of phenolic groups of six phenolic acid molecules with a FeCl3 molecule.
Gallic acid readily reacts with iron(III) chloride and forms a blue-black reaction product (ink).
Nucleophilic substitution reaction with halogens
The presence of phenolic groups (-OH) in the molecule of phenolic acids allows them to enter into reactions of substitution of hydrogen atoms of the benzene ring with halogens at normal conditions. Benzoic acid and ordinary aromatic acids do not enter into such reactions.
For example, bromination of salicylic acid:
HOC6H4COOH + Br2 → HO(Br)C6H3COOH + HOC6H3(Br)COOH + 2HBr salicylic acid p-bromosalicylic acid o-bromosalicylic acid
The phenolic group in the salicylic acid molecule acts as a substituent of the first kind - it directs atomic groups and individual atoms to replace the benzene ring with a hydrogen atom in the o- and p-position relative to itself.
Typical representatives and their derivatives, the use of phenolic acids in medicine and industry
2-hydroxybenzoic or salicylic acid is a typical phenolic acid, it is sometimes called hydroxybenzoic acid HOC6H4COOH. Salts and esters of salicylic acid - salicylates. It was first extracted from esters contained in the tissues of some plants - wintergreen and essential oil of Gaultheria procumbers. Salicylic acid is a crystalline solid. Has a bactericidal effect. Its salts and esters are widely used in medicine and veterinary medicine as medications. Salicylic acid is widely used in the production medicines(for example, acetylsalicylic acid, phenyl salicylate), mordant dyes, fungicides (for example, salicylanilide), odorous substances (methyl salicylate, benzyl salicylate), antiseptics in the food industry, in canning, as a reagent for colorimetric determination in solutions of iron and copper, as an acid-base indicator for luminescent analysis (at pH = 2.5...4.6 and in the presence of acid, blue luminescence appears), etc.
3,4,5 - Trihydroxybenzoic, or gallic acid - a phenolic acid containing one
Read also:
|
The herb and flowers of meadowsweet are used for medicinal purposes. The leaves, flowers and roots of meadowsweet contain flavonoids, phytoncides, steroids, terpenes, glycosides, tannins, starch, essential oil, vanillin, and, most importantly, large quantities, vitamin C. The meadowsweet herb contains 0.2% essential oil (the components of which are methyl salicylate and its bioside gaultherin, salicylic aldehyde, vanillin), phenolic glycosides (monotroposide, primveroside, salicin, spirein), flavonoids (4%: avicularin, hyperoside , spireoside), tannins - 15%, coumarins, steroids, carotene, vitamin C, trace elements.
Let's take a closer look at the action and properties of chemical compounds of meadowsweet.
Vitamin C. Participates in the biosynthesis of corticosteroid hormones, which are responsible for the adaptive reactions of the body; increases immunity; eliminates inflammation; stabilizing effect on connective tissue; improves the psycho-emotional state because it affects the formation of dopamine, norepinephrine, as well as serotonin and endorphins.
Tannins. These substances precipitate protoplasmic proteins, thereby exerting either an irritating or astringent effect on the mucous membranes (it all depends on the concentration of the solution). Properties: astringent; hemostatic; antioxidant.
Phenolic compounds (monotropitin, spirein). Properties: stimulating (activate the functioning of the adrenal cortex); antiseptic; diuretic; adaptogenic (increase protective forces organism); antispasmodic; sedative; choleretic; hemostatic.
Salicylic acid. Action: normalizes capillary permeability, thereby preventing tissue swelling; participates in the formation of inflammatory mediators; suspends the biosynthesis of prostaglandins, which play an important role in the development of inflammation, as well as pain, which relieves inflammation.
Figure 5.1 – Salicylic acid
Flavonoids: avicularin. Properties: anti-inflammatory; antiallergic; antivirus; anti-carcinogenic; antioxidant; choleretic; antiulcer; diuretic; antispasmodic.

Figure 5.2– Avicularin
Catechins. These substances neutralize free radicals, thereby preventing the development of cancer. In addition, catechins resist the effects of bacteria and prevent cell destruction, thereby significantly slowing down the aging process of the body.
Phenolcarboxylic acids. Action: relieve inflammation; accelerate the process of bile excretion; enhance kidney function; stimulate the antitoxic function of the liver.
Essential oils. Ingredients: methyl salicylate and salicin. Effect of essential oils: normalize functioning of cardio-vascular system; soften cough; enhance the separation of mucus from the bronchi; improve the functioning of the gastrointestinal tract. Properties: bactericidal; anti-inflammatory; antiseptic; stimulating; sedative.
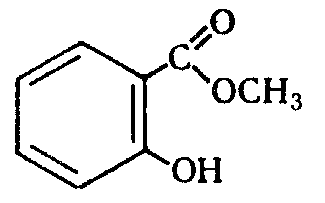

Figure 5.3 – Methyl salicylate Figure 5.4 – Salicin
Fatty acid. Action of fatty acids: participate in the process of energy formation; participate in the construction of membranes, which make up the skeleton of cells; normalize metabolism, being components of various lipids.
Glycosides. Properties: diuretic; antimicrobial; sedative; laxative; expectorant; vasodilator; disinfectant. Starch. Used as an enveloping agent in the treatment of inflammatory gastrointestinal diseases. In addition, starch belongs to the class of easily digestible carbohydrates, which, when transformed into glucose, quickly saturate the body with energy.

Figure 5.5 – Starch
Wax. It has bactericidal and astringent properties, therefore it is used in the treatment of difficult-to-heal wounds and various skin diseases.
In 1828, the Munich pharmacist Johann Büchner isolated the anti-inflammatory substance salicin from willow bark, the hydrolysis of which produced salicylic acid, and in 1838 the Italian chemist R. Piria isolated spireic acid from meadowsweet (Spiraea vinosa), which turned out to be a derivative of salicylic acid. It turned out that there is a lot of salicylic acid in the meadowsweet - Spiraea salicifolia, and then Felix Hofmann, an employee of the German company Bayer, developed a technology for the organic synthesis of acetylsalicylic acid, which received the commercial name “aspirin”. This name is made up of two parts: “a” from acetyl and “spira” from Spiraea. In meadowsweet, aspirin is in glycosylated form. More recently, diterpene alkaloids, called spiramins and spiratins, have been isolated from the seeds and roots of meadowsweet. Their action is similar to camphor, caffeine, but their use does not cause an increase in blood pressure. Scientists believe that spiramins protect brain cells from oxygen starvation. Substances that prevent the formation of blood clots have been isolated from Japanese meadowsweet.
The tannins of meadowsweet, undergoing oxidation, turn into substances with a bloody color - phlobafens .
| | 3 | | | | | | | |
In addition, they are characterized by properties due to the presence in the molecule of both types of functional groups and a benzene ring.
Phenol acids are crystalline solids. Phenolic acids, which contain one phenolic hydroxyl, are relatively poorly soluble in cold water, but dissolve well in hot water and many organic solvents. With an increase in the number of phenolic hydroxyls, the solubility of phenolic acids increases.
Distribution in nature[ | ]
Phenolic acids are very common in nature, so they can be extracted from natural raw materials (such as blood red hawthorn, chokeberry, propolis). Phenolic acids are the main component (55-85%) of the residue from the distillation of wood resin from wood pitch.
Synthesis [ | ]
Synthetic methods are often used to obtain phenolic acids. In particular, 2-hydroxybenzoic (salicylic) acid is extracted from carbon dioxide in autoclaves at 180 °C, followed by treatment of the reaction product with hydrochloric acid, Kolbe synthesis:
C6H5ONa + С02 180 С → С6Н4 (ONa) COOH; C6H4 (ONa) COOH + HC1 → C6H4 (OH) COOH + NaCl
Chemical properties[ | ]
Phenol acids simultaneously have the properties of carboxylic acids and phenols. In addition, they are characterized by properties due to the presence of both types of functional groups and a benzene ring in the molecule.
Heat decomposition[ | ]
When heated, phenolic acids decompose to form phenolic compounds and carbon dioxide. For example, when heated, salicylic acid decomposes into phenol and carbon dioxide:
HOC6H4COOH → C6H5OH + CO2 salicylic acid phenol carbon dioxide
Esterification reaction (at the carboxyl group)[ | ]
Phenol acids, like carboxylic acids, are capable of forming esters due to the presence of carboxyl and hydroxyl groups.
For example, the formation of salicylic acid ester - acetylsalicylic acid:
HOC6H4COOH + H3C-C(=O)-O-(O=)C-CH3 → C6H4(COOH)-O-CO-CH3 + CH3COOH salicylic acid acetic ester acetylsalicylic acid acetic acid
and the formation of gallic acid ester, one gallic acid molecule reacts with its carboxyl, the other with its phenyl hydroxyl. Chinese tannin is a glucoside of digallic acid and glucose.
Formation of salts[ | ]
Phenol acids, like carboxylic acids, form salts. For example, the formation of sodium salicylate by the interaction of salicylic acid and sodium hydroxide:
HOC6H4COOH + NaOH → HOC6H4COONa + H2O salicylic acid sodium salicylate
Reaction with iron(III) chloride (at the phenolic group)[ | ]
There is often a need to detect the presence of salicylic acid and other phenolic acids in canned foods. Then 2-3 ml of the test solution is placed in a test tube and a few drops of a 1% solution of iron (III) chloride are added. A purple color appears. Unlike phenol, it can also appear in an alcohol solution of acid. The color occurs as a result of the formation of complex salts during the interaction of phenolic groups of six phenolic acid molecules with a FeCl3 molecule.
Gallic acid readily reacts with iron(III) chloride and forms a blue-black reaction product (ink).
Nucleophilic substitution reaction with halogens[ | ]
The presence of phenolic groups (-OH) in the molecule of phenolic acids allows them to enter into reactions of substitution of hydrogen atoms of the benzene ring with halogens under normal conditions. Benzoic acid and ordinary aromatic acids do not enter into such reactions.
For example, bromination of salicylic acid:
HOC6H4COOH + Br2 → HO(Br)C6H3COOH + HOC6H3(Br)COOH + 2HBr salicylic acid p-bromosalicylic acid o-bromosalicylic acid
The phenolic group in the salicylic acid molecule acts as a substituent of the first kind - it directs atomic groups and individual atoms to replace the benzene ring with a hydrogen atom in the o- and p-position relative to itself.
Typical representatives and their derivatives, the use of phenolic acids in medicine and industry[ | ]
2-hydroxybenzoic or salicylic acid is a typical phenolic acid, it is sometimes called hydroxybenzoic acid HOC6H4COOH. Salts and esters of salicylic acid - salicylates. It was first extracted from esters contained in the tissues of some plants - wintergreen and essential oil of Gaultheria procumbers. Salicylic acid is a crystalline solid. Has a bactericidal effect. Its salts and esters are widely used in medicine and veterinary medicine as medicines. Salicylic acid is widely used for the production of medicines (for example, acetylsalicylic acid, phenyl salicylate), mordant dyes, fungicides (for example), odorants (methyl salicylate, benzyl salicylate), antiseptics in the food industry, in canning, as a reagent for colorimetric determination in iron solutions and copper, as an acid-base indicator in luminescent analysis (at pH = 2.5...4.6 and in the presence of acid, blue luminescence appears), etc.
3,4,5 - Trihydroxybenzoic, or gallic acid - a phenolic acid containing one
MINISTRY OF AGRICULTURE. USSR
ALL-UNION RESEARCH INSTITUTE OF PHYTOPATHOLOGY
- : ; - ; : " " ;
The manuscript is written by Lydia Vladimirovna ROZUM. ■ ■ 4 .
PHENOLCDRBONIC ACIDS OF WHEAT PLANTS DAMAGED BY STEM RUST
(01/06/11 - phytopathology and plant protection)
dissertations for the degree of candidate of biological sciences ■
MOSCOW -1977
The work was carried out in the laboratory of biochemistry and plant physiology ■ North Caucasus Research Institute of Phytopathology. * -
Scientific supervisors: Doctor of Biological Sciences, Professor M. I. Zaprometov, Candidate of Agricultural Sciences V. V. Chigrii.
Official opponents: Doctor of Biological Sciences, Professor V., I. Kefeli, Candidate of Biological Sciences V..V. Ma-
Leading institution ~ All-Union Research Institute of Plant Protection.
The dissertation defense will take place on 197
at 10 o'clock at a meeting of the specialized Council of the All-Union Scientific Research Institute of Phytopathology,
The dissertation can be found in the institute's library. , ""
Scientific Secretary- ;
candidate of agricultural
Sciences; . ■ ^
G. V. Pyzhnkova
GENERAL DESCRIPTION OF WORK
Relevance of the problem. Currently, crop protection from pests and diseases is becoming more important. One of the harmful diseases of grain crops is wheat stem rust, which causes huge yield losses and reduces the commercial quality of grain (Peterson, 1958; Chumakov et al., 1965). The main way to combat this disease is to develop resistant varieties. However, despite undoubted successes, breeding measures alone cannot lead to a radical solution to this problem, since sooner or later biotypes of the pathogen become widespread in nature and can infect new varieties. The advances achieved so far in the field of pathophysiology suggest that identifying the physiological and biochemical mechanisms of resistance to the pathogen will contribute to targeted research effective means protecting plants from rust.
Information about changes in phenolic metabolism in wheat plants when infected with rust is very scarce and rather contradictory. Analysis of literature data shows that the lack of a common point of view on the role of phenolic compounds in latogy "¡¿¿^¿^1 i.aV.^TT%VL[sledD°"
MOPiSH" Li., l. "l t i"V. I 1
vateln determined either the total content of all polyphenols in healthy and infected plants (Kiraly and Farkas, 1962; Seevers and Daly, 1970), or the amount of one particular metabolite (El-Naghy, 1963, etc.). In addition, these studies were carried out, as a rule, on very young plants. In nature, older plants are exposed to a real threat of rust infection, and with age, the mechanism of response reactions can change significantly (Chern et al., 1969; Bateman et al., 1965).
Purpose and objectives of the study. The purpose of this work was to study the metabolism of phenol to rbo acids in wheat plants when infected with stem rust. Phenol-carboxylic acids are key metabolites of phenolic metabolism and are precursors of a number of aromatic compounds and lignin (Zanrometov, -1964, 197Ü; Towers, 1968). They have high physiological activity and are involved in a wide variety of metabolic processes, and there is reason to believe that they play an important role in the formation of hypersensitivity.”
The objectives of the study included:
1) study of post-infectious changes in the main phenol carboxylic acids of wheat at the beginning of the development of the disease and at the time of the appearance of visible symptoms of the disease;
2) determination of the activity of enzymes related to the accumulation of these phenolcarboic acids;
3) obtaining information about the fungal and cytotoxicity of phenolic compounds of wheat, about their physiological functions.
Scientific novelty, practical value of the work and ways of implementation. For the first time, a systematic study of the metabolism of phenolcarboic acids in wheat plants affected by stem rust was carried out. The earliest changes in the content of phenolcarboic acids during infection, which are directly related to necrobiosis, were studied. It was found that the development of protective necrogenic reactions is preceded by the accumulation in the tissues of resistant plants of certain phenolcarboic acids, which have high fungonic and cytotoxicity. In contrast to the well-known works on the role of polyphenols in the resistance of wheat plants to infectious diseases, our research was carried out on adult wheat plants, which in nature are exposed to a real threat of rust infection. The experimental results are important for understanding the mechanisms of plant defense reactions and can be used in the targeted synthesis of new anti-rust fungicides.
Approbation. The dissertation materials were presented at the All-Union Symposium on Phenolic Compounds (Alma-2
Ata, 1971) and at the Third All-Union Symposium on Phenolic Compounds (Tbilisi, 1976).
Workload. The dissertation is presented on 137 pages of typewritten text and consists of an introduction, three chapters of the experimental part, a conclusion, a list of cited literature; includes 12 figures and 21 tables in the text. The list of used literary sources contains 230 titles, of which 131 are in foreign languages.
Section I of the review provides data on the wide distribution of phenolic compounds in the plant world and lists the most important functions of these compounds in metabolic processes. Section II reveals the important role of polyphenols in plant resistance to fungal, bacterial and viral diseases.
EXPERIMENTAL PART Chapter!. MATERIALS AND METHODS OF RESEARCH
The studies were carried out on two pairs of varieties: Kaplya and Emmer, belonging to the species Triticum dicoccum, Balladi 116 and Kubanka 3, belonging to the species Triticum durum. Plants were grown in a greenhouse until the booting phase and infected with freshly collected uredospores of the 40th race of Puccinia graminis Pers f. sp. tritici Eriks, et Herrn. The first variety of each pair is resistant to this race of rust, the second is susceptible. When the variety Balladi 116 was infected, the reaction type was 0-0; Drops-0; -1, Emmer and Kubanka 3-3-4 according to Goyman’s classification (Goyman E., 1954). Thus, the varieties Kapl n Balladi 116 were an incompatible combination with this pathogen, while the varieties Emmer and Kubanka 3 were compatible.
Samples of infected and control (uninfected) plants were taken at the very beginning of pathogenesis (the second day after infection) and during the reproductive phase of fungal development (the fifth day). Whole leaves were frozen with liquid nitrogen, ground, lyophilized, sealed in ampoules and kept cold for analysis.
Soluble phenolic compounds were extracted with hot 80% ethanol. The latter was evaporated, the aqueous residue was acidified to pH 2.0, and free phenolcarboxylic acids were extracted with diethyl ether.
nolcarboxylic acids were determined after twenty hours of hydrolysis of the aqueous residue with a p-glucosidase preparation (0.07 mg/ml) at 37° C and subsequent extraction with diethyl ether. Soluble esterified forms of phenolcarboxylic acids were determined after five hours of hydrolysis (2 ppm NaOH in nitrogen atmosphere) of the aqueous eluate remaining after treatment of the solution with p-glucosnase. The bleached plant material remaining after the extraction of soluble polyphenols was incubated for five hours in a nitrogen atmosphere with 2 and NaOH, and the ethanol-insoluble ether-linked phenolcaboxylic acids released during hydrolysis were extracted with diethyl ether after acidification of the mixture to pH 2.0, Separation of phenolcarboxylic acids was carried out by two-dimensional chromatography using 1% acetic acid in the first direction and the organic phase of a mixture of toluene-acetic acid - water (4:1:5) in the second direction.The chromatograms were treated with a weak solution salts of strong red GG, phenolcarSyunovys acids were eluted from paper with 0.05 N. "NaOH in ethanol and used for quantification.
Determination of glycosidase activity was carried out on protein preparations isolated from healthy and infected plants, using the corresponding l-nitrophenol glycosides as substrates (Jaynes et al., 1972).
Phenylalanine-lmmonia lyase activity was determined by the method of Koufcol and Conn (1961).
The ability of wheat plants to bind phenolic compounds was studied using the example of ferulic acid, which was introduced into plants in the form of a 2-10~3 M solution by absorption through leaf sections. The intensity of binding was judged by comparing the amount of acid remaining in the free state with the amount found in hydrolyzed forms 3 hours after the end of solution absorption.
The effect of phenolic compounds on germinating uredo-spores of stem rust was studied in the concentration range from 10 g M to 2.5 10-5 M. When studying the effect of various phenolic compounds on the toxicity of ferulic acid, the latter was used in concentrations of 2-10-3 M and 5 lO-4 M. Other phenolic compounds were added to ferulic acid in ratios of 1:5; 1:2; 2:1; 5:1. Solutions of fepol compounds were mixed with agar, applied to glass slides, and uredosiors were sprayed onto them. Aged glass-4
18 hours in the dark at 18-20° C. Counting of germinated spores was carried out in 10 fields of view of the microscope.
The cytotoxic properties of feiolcarboxylic acids were judged by the breakdown of chlorophyll in sections of wheat leaves incubated in solutions of the studied compounds. Determination of chlorophyll content was carried out according to the method described by Kulaeva (Kulaeva, 1973). In addition, the cytotoxic properties of phenolic compounds were assessed by their effect on the ability of leaf tissues to absorb acid fuchsin dye from solution; the dye was then eluted from the tissues and colorimeterized at 545 nm.
To study the possibility of increasing the resistance of wheat plants to rust (by increasing the level of endogenous polyphenols), infected plants of the susceptible variety Kubanka 3 were treated three times with solutions of phenol carboxylic acids in a concentration of 10-3 M. On the ninth day, the number of developed pustules on control and treated plants was taken into account.
The results of the experiments were subjected to variational statistical processing using Student’s test (Snedekor, 1961) at a probability level of P = 0.95,
Chapter II. POST-INFECTION CHANGES IN THE CONTENT OF PHENOLCARBOXYLIC ACIDS
AND LIGNIN IN WHEAT PLANTS WHEN INFECTED BY STEM RUST
Phenolcarboxylic acids (PCAs) of wheat plants are found both in the free state and in the composition of glycosides and esters, both soluble in ethanol and insoluble. The total amount of all forms of PCA in wheat leaves ranged from 2228-3259 μg/g of dry matter, "The content of soluble esters averaged 49% of the total amount, insoluble - 39%, glycosides - 9% and free PCA - 3%. In In the fractions of free, glycoside-bound, ester-bound soluble forms of FCA, you identified ferulic, vanillic, lilac, p-coumaric, p-hydroxybenzoic acids, while in the fraction of insoluble esters - only ferulic, vanillic and p-coumaric acids. , are present in all FCA fractions, almost always predominate quantitatively, and can probably be considered one of the main components of the phenolic complex of wheat plants.
Infection of wheat with stem rust causes significant changes in the metabolism of PCA. Table 1 data
(% to control)
Phenolcarboxylic ACIDS Incompatible combination Compatible combination
Ka pli Ba.plali 116 Emmer Kubanka 3
Ferulic...... 233* 171* 64* 85
Vanillin..... 183* 125 76 10)
Lilac...... 126* 117 194* 85
p-Kumaronaya..... 169* 131* 127 100
p-OxybeshoAiaya.... 133 120
* Differences are significant at P=0.95.
indicate that resistant varieties are characterized by the accumulation of free fepolcarboxylic acids, in contrast to susceptible varieties that do not have this ability. In the resistant variety Drops, which gives a typical “hypersensitivity” reaction, at the beginning of the development of necrobiosis, the content of all FCAs significantly increased. The content of ferulic acid, which is the most toxic to the pathogen, in this case more than doubles. In the Balladi 116 variety, whose reaction is closer to the immune one, an increase in all phenol carbonic acids was also found. Vanillic and syringic acids accumulate, which can enhance the toxic effect of ferulic acid. In susceptible varieties, rust infection, as a rule, was not accompanied by the accumulation of free fepolcarboxylic acids. Moreover, in the tissues of the Emmer and Kubanka varieties, the content of ferulic acid even decreased, and the amount of other FCA also decreased or remained at the control level. The noted changes persisted on the fifth day after infection, however, they are not so clearly expressed and, apparently, are less important from the point of view of elucidating the factors of necrobiosis.
Infection did not cause significant changes in the glycoside fraction, although a certain tendency towards a decrease in their level can be noted by the fifth day after inoculation, especially in resistant varieties. Significant changes were observed in the fractions of soluble and insoluble esters. As the data in Table 2 show, the content of ethanol-soluble FCA esters decreased sharply already on the second day after infection. This decrease is especially significant in the resistant variety Kaplya, which is characterized by b
Phe i i o lka r bo n oic acids Incompatible combination Compatible combination
Drops Ballady 116 Emmer Cuba Yak 3
days after infection.
2 5 2 5 2 5 2 i-
Ferulic...... 37* 36* 91* 90* 68* 70* 99 98
Vanillin.....zo* 36* 64* 78* 37* 79 115 98
Lilac...... 10* 38* 79* 77* 36* 108 89 96
i-Kumarovaya..... 28* 10* 102 87 33* 66* 69* 88
l-Oxybnzoylaya. . . . 97 73* W 114
* Differences are significant at P = 0.95,
extensive microbiosis at the sites of pathogen entry. The content of ferulovone, vanillic, and syringic acids also decreased in the resistant variety Ballad" 116. In the susceptible variety Emmer, infection also contributes to a decrease in ester-bound ethanol-soluble PCA, but these changes are less pronounced. In the highly susceptible variety Kubanka 3, the amount of soluble esters practically does not change under the influence of infection. "
Quite clear differences between resistant and susceptible varieties are found in the fraction of insoluble esters associated with cell wall material (Table 3). Resistant varieties respond to infection by a sharp decrease in the content of essential ferulic acid. This fact may indicate that some reactions in the metabolism of this acid play an important role in the process of pathogenesis. The amount of vanillic and p-coumaric acids also tends to decrease in resistant plants at the beginning of disease development. In contrast, in susceptible plants, infection, as a rule, does not cause a decrease in the content of insoluble forms of PCA, and in some cases a noticeable increase can even be observed.
Quantitative change data various forms FCCs indicate that infection has different effects on the phenolic metabolism of resistant and susceptible wheat plants. Thus, resistant varieties are characterized by a significant accumulation of free PCA, which have a high
Phe ol carbo noic acids Incompatible combination Compatible combination
Drops Balladi 11 b Emmer Kubanka 3
days after infection
Ferulic 36* 25"" 27* 36* 98 128* 80 152*
Vanillin 74* 115 85 103 167* 99 166* 127*
p-Kumarovaya 83* PZ 75 101 I 83* 90 26* 124
* Differences with control are significant at P = 0.95.
some physiological activity (Bardinskaya et al., 1962; Kays et al. 1956), and quite profound changes in the fraction of bound forms of phenolic compounds. In contrast, in susceptible varieties, only the amount of ethanol-soluble forms of PCA undergoes some changes. At the same time, ethanol-insoluble FCCs are not noticeably affected.
It seemed likely that the accumulation of free PCA could occur due to: a) an increase in the activity of hydrolytic enzymes, among which the most interesting are glycondases, which are capable of hydrolyzing not only glycoeides, but also some esters (Towers, 1968); b) enhancing de novo biosynthesis of FCA and c) suppressing the processes of their binding in the form of glycosides, esters and lignin (Towers, 1968). The results of determining glycosidase activity, presented in Table 4, show that in the immune variety Balla-dn 116, both p- and a-glycosidases are activated; in the resistant variety Drops, which forms extensive necrosis, only p-glycosidases are activated. Glycosidase activity of susceptible plants, as a rule, decreases or remains at the control level.
A study of the activity of phenylal anine ammonia lyase (NF 4.3.1.5) - (PAL-lyase), which plays an extremely important role in the biosynthesis of phenolic compounds, showed that in a resistant variety, infection leads to a significant increase in the activity of this enzyme (Fig. 1) . In the susceptible variety, a slight increase in PAL-lyase activity or even a decrease in its activity was observed compared to the control, especially in initial stage development of infection.
days after JTT
Rice. І Akpmosti fshlshdon - shsh jasper b tshtshsh litih doshtsm.
Determination of the intensity of binding of exogenous ferulic acid showed that at the beginning of pathogenesis in the tissues of uninfected leaves of the Kaplya variety, 98% of this compound was converted into bound forms in 3 hours, while in infected leaves - only 95%. In the Emmer variety, the binding intensity during infection increased from 93 to 98%. Although these differences are small, they are statistically significant and indicate a weakening of the binding processes of phenolcarboxylic acids when a resistant variety is infected and an increase in the susceptible variety.
Table 4
Glycosidase activity (mg substrate/m(Gprotein/hour) in Wheat leaves (two days after infection with stem rust)
Glancosidases Incompatible combination Compatible combination
Drops Balladn 116 Emmer Kubanka 3
healthy infected healthy infected healthy infected healthy infected
P-glucose ida for 338 389 75)E>9 508 387 595 496
a-glucose and yes for 6 6 0 7 10 5 7 8
p-galactosidase 165 183 46 130 273 226 101 102
a-galactoside az a 89 85 39 99 51 37 46 31
resistant (Kapli) varieties, suggests that in the first case, the success of the protective reaction is associated not so much with the amount of released PCA, but with the speed of this process, due to the sharp activation of p- and a-glycosidases. Thus, the accumulation of free PCA in the leaves of a resistant variety may be associated with increased glycosidase activity of leaf tissues, activation of PAL-lyase and weakening of free acid binding processes.
As mentioned above, the proportion of soluble and insoluble FCA esters amounts to a total of 88% of the total amount of wheat FCA, while the share of free ones accounts for 3%. "It is obvious that the post-infectious loss of PCA esters (Tables 2 and 3) cannot be explained solely by an increase in their hydrolysis, which may be partially associated with the accumulation of free PCA in the leaves of resistant plants (Table 1). The question arises: possible reasons such a sharp decrease in esterified PCA, especially noticeable in the leaves of resistant plants. One of these “reasons is probably the intensification of respiration of infected plants (especially resistant ones) and the associated large loss of carbon in the form of CO2. The study of this issue showed that on the 2nd day after infection in the varieties Kapli and Emmer, the release of CO2 from the leaves increased by 54% and 45%, respectively.A sharp increase in carbon losses could well reduce the volume of biosynthesis of secondary compounds (including lignin) due to the need to maintain the concentrations of biologically more important metabolites at an optimal level.
Another reason for the loss of PCA esters in leaves may be the redistribution of phenolcarboxylic acids between above-ground organs and roots. One confirmation of the validity of this assumption is the fact that in the roots of plants of the resistant variety Drops at the beginning of the disease, the amount of free PCA during infection increased by 43%, bound (soluble and insoluble) by 78%, and the total amount by 75%. In the Emmer variety, only the content of the first fraction increased noticeably (by 58%). Total FCC, including related forms, increased slightly (by 10% and 7%, respectively). Thus, comparing the data in Tables 2 and 3 with the results of determining PCA in the roots, it can be assumed that the accumulation of phenolcarboxylic acids in these organs correlates to a certain extent with the loss of associated forms of PCA in the leaves. The data obtained are not enough to provide a complete interpretation of the relationship between the phenolic metabolism of above-ground organs and the root system. It is possible that the decrease in PKK esters in infected leaves is due to the fact that infection can cause changes in metabolism in organs distant from the site of pathogen development (Pac|ecb, 1974), and increased biosynthesis of PKK in the roots leads to the “interception” of some precursors , which are normally directed from the roots to the leaves. Apparently, we cannot exclude the possibility of the outflow of directly soluble PCA from the leaves to the roots. Our experiment showed that when wheat roots are immersed in solutions of 4-methyllumbellifero-]0
countertmr M
I<г* uf* w* mi* b"ti3 /гг
Fmomya -acid-"
II "spherical acid
".........- bscinic acid
Puc. h Toxicity of phenolic acids
No. OF CELLS OF MESTENNY PSHESCHY.
On n-hydroxybenzoic acid, these substances accumulated in leaves and stems.
Thus, a sharp decrease in PCA esters in infected leaves of resistant varieties may be associated, on the one hand, with a pathologically increased respiration rate and, on the other, with an increase in PCA biosynthesis in the roots.
Chapter III. STUDYING THE FUNGI- AND CYTOTOXICITY OF PHENOLIC COMPOUNDS
Ferulic acid and coniferyl alcohol are the most toxic to germinating fungus uredoslors, which, even at a concentration of 5* 10-" M, suppressed spore germination by 44% and 38%, respectively. Other phenolic compounds found in the plant cell can have a varied effect on toxicity quantitatively predominant ferulic acid.Indeed, the data presented in table 5 allow us to note that.
Table 5
Toxicity of phenolic compounds and their mixtures to rarely germinating stem rust ospores of wheat
Compound % suppression of uredoslor germination under the influence
substances in a concentration of 5 ■ KMM material ti (M) mixed with ferulic acid 5-10-1 M
2.5 > I0-g (5:1) 10-" (2:1) 5- 10"* (1: 1) 10-" (1:5)
Ferulic acid. . . 44
Conifer alcohol 33 77* 74* 76* 50
Vannlil..... 0 100* 71* _ 67*
Vanillic acid. . 0 95* 75* 55* 27*
Lilac acid. . 0 60* 45 49 59
Protocatechuic acid 0 - 19* 16* 13*
Quercetin...... 50 29 15* 28 32
* Distinguished between this option and the pure one! ferulic acid are significant at P = 0.95.
for example, conferyl alcohol showed additivity to ferulic acid. When adding other compounds, "mainly those having 4-hydroxy, 3-methoxy or 4-hydroxy, 3,5-dimethoxy groups (vanillic and syringic acid, vanillin) in the benzene ring), a certain synergism with ferulic acid is noted. At the same time, polyphenols with 3,4-dioxy groups (protocatechuic acid, quercetin) showed antagonism. Thus,
The nature of the toxic effect of the phenolic complex is very complex: it is determined not only by the concentration of the most toxic substances, but also by their ratio with substances that can enhance or weaken their effect.
Due to the fact that phenolic compounds, as it turned out, have a fairly high fungal toxicity, it seemed important to study the possibility of increasing the resistance of wheat plants by directly introducing individual phenolcarboxylic acids and their mixtures into leaf tissue.
The data presented in Table 6 allows us to note that treatment of leaves of a susceptible variety with FCA solutions leads to a decrease not only in the degree of damage, but also to a simultaneous change in the type of plant damage. Thus, if in the control variant the type of lesion was 3-4 points, then in the variant with the participation of ferulic acid, and especially its mixtures with vanillic and syringic acids, the type of lesion was close to 1. The data in Table 6 are in good agreement with the idea that A post-infectious increase in the concentration of FCA in leaves may be one of the factors responsible for the incompatibility of partners.
Phenolic compounds, accumulating in the zone of pathogen penetration, can apparently also exhibit a cytotoxic effect. Since the first obvious symptom of a “hypersensitivity” reaction is the breakdown of chlorophyll
*Table 6
Changes in the response of wheat plants to stem rust infection under the influence of their treatment with 10-3 M solutions of feriolic acids
Intensity
Infection experience options Type
number % to reaction control
pustules per leaf *
Control (water)....... 16 100 3-4
Ferulic acid...... 10 63 1-2
Vanillic acid.... 9 56 1-2
Lilac acid...... II 69 2
Protocatechuic acid, . . 13 81 3
p-coumaric acid..... . 13 81 2-3
p-hydroxybenzoic acid.... 1! 69 2-3
Ferulic + protocatechuic acid
you............ 13 81 .2
Ferulic + Vaillin Yasirene- 43
high acid........ 7 1
Ferulic + new vanilla + - lilac - 56 1
va i + protocatechuic acid 9
* The smallest significant difference at P=0.95 is 3.
^__o o*-- 1 40
whether to "* ">"s w3 »w"»
"- Lol deііstshi ferulovo" kshshtm.
*-- product is found.
Rice. Items of fershmi acid h product ft oxidation
la in the infection zone and the associated formation of a chlorotic spot, it could be assumed that the change in the amount of chlorophyll in leaf segments incubated in FCA solutions serves as a measure of the toxicity of polyphenols for plant cells. Our study of the cytotoxicity of phenolcarboxylic acids showed that ferulic and syringic acids at concentrations of 4-10"3 M and 6-10~3 M caused the destruction of chlorophyll in sections placed on solutions of these acids. In other experiments on the cytotoxic effect of FCA were judged by the ability of the plant tissues they treated to absorb the dye - acid fuchsin - from the solution. The herbicide paraquat with a clearly expressed necrogenic effect on plant tissues was used as a toxicity standard. Glucose and sodium chloride were chosen as obviously non-toxic substances (in the concentrations we used). The data presented in Figure 3 allow us to conclude that phenolcarboxylic acids exhibited a cytotoxic effect and in this regard they formed the following series: ferulic>p-coumaric>vanillic.
However, in plants in vivo, phenolic compounds can be even more toxic. It is known that under the action of phenol-oxidizing enzymes, polyphenols are converted into quinones (Stom, 1972, 1973), which are significantly more toxic than the original phenolic compounds (Stom, 1970). We studied the effect of the oxidation product of ferulic acid (transformation of the latter was carried out by horseradish peroxidase for 30 minutes in the presence of HgO2) on sections of wheat leaves. The data presented in Figure 4 suggests that the oxidation product of ferulic acid had more high cytotoxicity compared to the original form of this compound.
Thus, phenolcarboxylic acids, and also, probably, their oxidation products, can exhibit toxicity both to the pathogen and to the plant cell itself.
Conclusion
We have established that already at the very beginning of the development of the stem rust pathogen in the tissues of resistant wheat plants, the activity of glucosidases and phenylalanine ammonia lyase increases, while the intensity of binding of phenol carboxylic acids in the form of soluble and insoluble esters and lignin, on the contrary, decreases. This leads to the accumulation of free phenolcarboxylic acids in infected leaves, among which ferulic acid predominates, which is characterized by noticeable fungi- and cytotoxicity.
This accumulation, of course, occurs locally at the sites where the pathogen comes into contact with the leaves of the plant, and this local increase in concentration is, in all likelihood, much more significant than what we found when analyzing the leaf as a whole. Indirect confirmation that the observed We associate changes in the phenolic complex of leaves with contact zones, is the fact that the development of extensive necrosis on the leaves of the Kaplya variety was preceded by a stronger increase in the concentration of free PCA than in the case of the immune variety Balladi 116 with its point necrosis.
Upon contact with the peroxidase system, the formation of oxidized products may occur, which are characterized by higher fungi- and cytotoxicity than the parent compounds. Such contact can occur as a result of post-infectious disturbances in the intracellular compartmentalization (Metlitsky, Ozertskovskaya, 1968). In any case, there is no doubt that when resistant wheat plants are infected with stem rust, the intensity of peroxidase oxidation of phenolic compounds increases sharply (Chigrip et al., 1969).
Our assumptions about the protective functions of phenolcarboxylic acids are also confirmed by the results of experiments in which susceptible plants were treated with solutions of these compounds. Naturally, we are far from connecting the development of a compatible or incompatible reaction to infection solely with the peculiarities of changes in the concentrations of certain phenolcarboxylic acids. Nevertheless, the data obtained in our work indicate that there is still a certain connection between early post-inoculation changes in the phenolic complex and the nature of reactions to stem rust infection.
The results of studying the characteristics of phenolic metabolism in wheat plants with different resistance to stem rust, carried out in 1968-1973, allow us to draw the following main conclusions.
1. Phenolcarboxylic acids of wheat leaves are present mainly in the form of ethanol-soluble and ethanol-insoluble esters, as well as in the form of glycosides and in the free state. The total amount of all these forms is 2.2-3.2 mg/g dry matter. Of this amount, the share of insoluble and soluble esters, glycosides and free acids accounts for an average of 39%, 49%. 9% and 3% and
respectively. In the last three fractions, ferulic, vanillic, lilac, p-coumaric and p-hydroxybenzoic acids were found, in ethanol-insoluble ferulic, vanillic and p-coumaric,
2. When resistant plants are infected with stem rust, already at the beginning of pathogenesis (2nd day after inoculation), the activity of glycosidases and phenylalanine ammonia lyase noticeably increases, while the ability of leaf tissues to convert free phenolcarboxylic acids into a bound state decreases. These changes lead to a sharp increase in the content of free phenolcarboxylic acids, especially ferulic acid. In susceptible plants, such changes were not detected; a slight increase in the content of phenolcarboxylic acids is observed by the 5th day after infection.
3. Infection of resistant plants is accompanied by a decrease in the content of bound phenolcarboxylic acids and lignin in the leaves. On the other hand, in susceptible plants, the content of only soluble phenolic compounds decreases, and the amount of insoluble esters of phenolcarboxylic acids even increases slightly,
4. Infection of wheat plants with stem rust uredospores also affects the phenolic metabolism of roots. At the same time, the changes occurring in the roots are opposite to those that are characteristic of infected leaves: the content of free phenolcarboxylic acids and their insoluble esters here increases significantly. These processes are more pronounced in resistant plants than in susceptible ones.
5. The high toxicity of ferulic acid for germinating uredospores of the pathogen has been established. Vanillic and syringic acids, being non-toxic to uredospores, joint use with ferulic acid significantly enhanced]! toxic effect of the latter. Ferulic acid also turned out to be the most toxic to wheat leaf tissue, with its oxidation occurring within 30 minutes. "peroxindase increased the toxicity (method of formation of an even more toxic product),
6. Treatment of infected susceptible plants with a solution of ferulic acid and especially its mixture with vanillic and lilac acid not only significantly reduced the intensity of infection, but also changed the type of reaction, bringing it closer to the type characteristic of resistant plants. The data obtained suggest that the local accumulation of phenolcarboxylic acids in the zone of pathogen development may be one of the reasons for cell necrosis and death of the pathogen in the tissues of resistant plants.
1. Changes in phenolic metabolism in spring wheat when infected with stem rust. Plant Physiology, 1969 (16), 2, 330-335 (co-authored with V.V. Chigrin).
2. Toxicity of phenolic compounds to germinating uredospores of wheat stem rust Puccinia graminis Pers f. sp. tritici Eriks, et Henn. Mycology and phytopathology, 1969, 3 (3), 243-248 (co-authored with V.V. Chigrin and L.M. Bessmeltseva).
3. Some features of phenolic metabolism in wheat varieties with different resistance to stem rust. Abstracts of the Second All-Union Symposium on Phenolic Compounds. Alma-Ata, 1970, 106 (co-authored with V.V. Chigrin).
4. Phenolcarboxylic acids and lignin in the leaves of resistant and susceptible varieties of spring wheat when infected with stem rust. Plant Physiology, 1973, 20, vysh, 5, 942-948 (co-authored with V. V. Chigrin, N. M. Zaprometov).
5. Some features of wheat phenolic metabolism and resistance to stem rust. Abstracts of the Third All-Union Symposium on Phenolic Compounds. Tbilisi, 1976 (co-authored with V.V. Chigrin, M, N. Zaprometov).
L 77690 1/1V-77 g. Volume ] p.l. Order 745.
Printing house of Moscow agricultural industry Academy named after K-A. Timiryazev 125008, Moscow A-8, Timiryazevskaya st., 44